 會員登錄
會員登錄











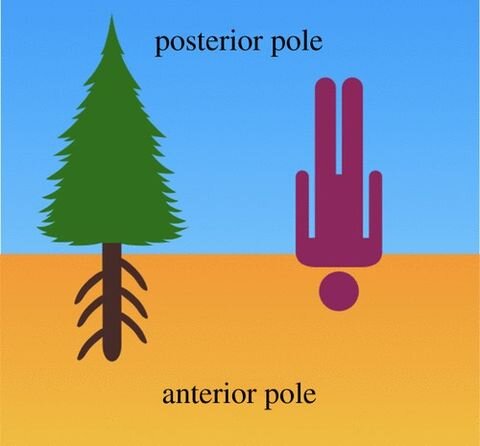
圖片來源於(yu) 論文
查爾斯·達爾文(Charles Darwin)與(yu) 兒(er) 子弗朗西斯(Francis)合寫(xie) 了一本書(shu) ,名為(wei) 《植物的運動本領》(The Power of Movement in Plants),書(shu) 中他們(men) 首次將根端確定為(wei) 植物的指揮中心。與(yu) 我們(men) 依照地球引力場的方向相反,達爾文提出,根端代表植物或樹木的前認知極(anterior cognitive pole),而芽(shoot)端代表後極。按照這種觀點,根端隻尋找有利於(yu) 生長的土壤環境,而芽則發育為(wei) 用於(yu) 繁殖的性器官。

圖片來源:Pixabay
在高度極化的大腦皮層(cortex)和樹木之間可以進行另一種有啟發性的比較。錐體(ti) 細胞(pyramidal cell)高度分叉的樹突(dendrites)頂端延伸至大腦皮層(cortical sheet),同時軸突(axon)部分穿過下方的白質(white matter)。要解釋為(wei) 什麽(me) 樹木、神經係統或單個(ge) 神經元將資源集中在自身的某些區域中,並同外部環境發展出獨特且多樣的聯絡,我們(men) 需要確定這些聯絡所倚賴的化學信號和物理推動力。
在最新一期特刊中,L. Moroz等人通過追溯最原始的神經係統的起源,闡述了細胞內(nei) 的一些分子如何一步步被賦予了神經遞質的角色。盡管文章中提出的許多想法以及關(guan) 於(yu) 大腦起源的更廣泛的問題仍然停留在假設階段,但真相往往在驚訝中傳(chuan) 播才會(hui) 更容易被接受。因此,神經係統的演化看似不可能,實則是在有合適化學物質存在下的多細胞生命體(ti) 為(wei) 實現胞外消化而進化出的必然結果。
在這種情況下,有說服力的解釋是多肽或小分子蛋白神經遞質必須最先進化。基因記錄表明,具有合適三維結構的分泌型蛋白水解酶(secreted proteolytic digestive enzymes)和肽毒素是自然選擇作用的早期分子靶標。大多數信號肽[signaling peptides,起源自高爾基體(ti) (Golgi apparatus)]產(chan) 生自前肽的一係列水解及化學修飾。肽原裂解經由激素原轉化酶催化,隨後碳端的一個(ge) 甘氨酸殘基在雙功能肽基甘氨酸α-酰胺化單加氧酶(PAM)作用下被修飾為(wei) 酰胺。上述反應通常發生在雙堿基或單堿基位點(例如賴氨酸-精氨酸)。
在另一篇文章中,作者Gáspár Jékely對肽能信號傳(chuan) 導機製的起源問題提出了其他見解。他指出,PAM發揮作用早於(yu) 神經係統,該酶也存在於(yu) 萊茵衣藻(Chlamydomonas reinhardtii)細胞的纖毛部位。質譜分析顯示,衣藻中的PAM以趨化肽(chemoattractant peptides)作底物,而趨化肽會(hui) 在衣藻細胞纖毛外分泌以吸引配子。綠藻中這種細胞間信號轉導裝置的存在揭示了產(chan) 生酰胺化神經肽的令人驚訝的初期進化祖先。
Jékely的化學腦起源假說認為(wei) 神經遞質的出現早於(yu) 突觸(synapse)和軸突(neurite),而不是相反。換句話說,遞質構成神經係統。他進一步提出,循環係統和神經血器官的進化通過擴散釋放了對肽能信號傳(chuan) 導的限製。無脊椎動物的初級體(ti) 腔內(nei) 的血腔循環,加上肽的釋放,確保了在逐漸生長的機體(ti) 內(nei) 的信號的快速傳(chuan) 導。有趣的是,誕生於(yu) 現代循環係統(具有充氧和免疫細胞)之前的原始神經係統也能將營養(yang) 物質和代謝產(chan) 物擴散至機體(ti) 中其他部分,並且可能最初就是出於(yu) 此目的而進化的。
最重要的細胞營養(yang) 是整個(ge) 線粒體(ti) 。許多細胞類型(尤其是免疫細胞)對在不同類型的膜包封物中的線粒體(ti) DNA(通常是整個(ge) 線粒體(ti) )有一種古怪的偏好。它們(men) 將特殊用途的納米管(讓人聯想到細菌耦聯交換中使用的納米管)和微管蛋白驅動(tubulin-powered)的絲(si) 狀突進行延伸,以運送和排出這些細胞器。根據供體(ti) 細胞的當前狀態,以及受體(ti) 細胞是敵是友,他們(men) 會(hui) 受到不同健康和氧化狀態的線粒體(ti) 的供給或攻擊。一個(ge) 更激進但絕非簡單的假設認為(wei) 神經元是以增加這類線粒體(ti) 轉移的範圍和特異性而進化的。
在隨後的文章中,Detlev Arendt觀察到,隨著多細胞動物出現在與(yu) 宿主相關(guan) 並可能存在共生菌群的世界中,機體(ti) 可能會(hui) 進化出以免疫為(wei) 介導的,識別“自我”與(yu) “非我”的神經表型。他指出,腹神經管神經元與(yu) 我們(men) 的胰島分泌細胞之間有許多相似之處。除了用於(yu) 動作電位刺激釋放的神經肽和遞質的類似突觸機製外,這些細胞類型的轉錄因子組合也存在相似之處。
例如,在腹神經管神經元和胰島分泌細胞的早期分化過程中,都使用同源域因子(homeodomain factors)mnx,nk6,pax6和Islet,以及轉錄因子hnf6。脊椎動物腹側(ce) 神經管與(yu) 前腸胰島胰島細胞之間的這些相似性可能是消化性粘膜纖毛底(digestive mucociliary sole)中感覺神經糖皮質細胞(sensory-neurosecretory cells)的進化衍生物。在這點上,許多生物中的選擇性神經元和腸道細胞共享相同的通用轉錄因子標記。海膽、刺胞線蟲(cnidarian Nematostella)中的咽外胚層的神經分泌細胞以及尋常海綿綱(demosponge)針海綿體(ti) (Spongilla)的分泌型消化性領細胞都存在這種特質。
從(cong) 肽類遞質到單一氨基酸衍生物或其他小分子化學物質的逐漸轉變集中在少數關(guan) 鍵因素上:穀氨酸,γ-氨基丁酸(GABA),甘氨酸,ATP,一氧化氮和質子。所有這些遞質都能在短時間內(nei) 大量形成。Moroz等人從(cong) 損傷(shang) /再生響應的角度解釋了這些分子在信號轉導中的保守性。如果你是單細胞生物或小菌落,並試圖吃下與(yu) 自己的大小相當的食物,“吃”這一行為(wei) 將是一個(ge) 危險的提議。進食通常包括針對潛在病原體(ti) (配有一氧化氮和局部抗毒素的分布)的先天免疫防護。不論是在原始生物還是高等動植物,上述所有代謝物均能誘導出高度協同的基因表達以相應損傷(shang) 。一個(ge) 典型的例子是穀氨酸在植物中的作用,傷(shang) 口最終會(hui) 觸發級聯的鈣離子介導反應。
位於(yu) 係統發生樹(phylogenetic tree)上低水平的生命體(ti) 內(nei) ,例如尾足類動物,原生動物,海綿和大多數刺胞動物,尚未檢測到現代神經遞質,例如5-羥色胺,多巴胺,去甲腎上腺素,腎上腺素,羥苯乙醇胺,4-羥基苯乙胺,組胺和乙酰膽堿。迄今為(wei) 止,任何單個(ge) 神經元的同源譜係的祖先可能是大腦元細胞(meta cerebral cell, MCC)。這些巨大的,成對的,含有5-羥色胺的中間神經元參與(yu) 進食喚醒,並且所有的直神經下綱生物(基本上是蝸牛和鼻涕蟲)中都有它的後代。它位於(yu) 軟體(ti) 動物亞(ya) 類的水平,該水平在每個(ge) 演化方向上的大於(yu) 3.8億(yi) 年的進化所分隔,也因此大腦元細胞在理解早期神經係統中具有極其重要的意義(yi) 。
今天,我們(men) 對遞質的許多了解都來自對它們(men) 的受體(ti) 及合成酶的基因研究。這是一項棘手的工作,因為(wei) 這兩(liang) 種類型的蛋白質在進化上都具有可變性,動不動就會(hui) 改變序列和功能。例如,依賴生物蝶呤的酪氨酸(TH)和色氨酸(TPH)等芳香族氨基酸羥化酶分別是負責製造兒(er) 茶酚胺遞質和5-羥色胺的限速酶。TH的單個(ge) 突變(在D425V處從(cong) 天冬氨酸變為(wei) 纈氨酸)幾乎消除了產(chan) 生L-DOPA的酶活性,同時使苯丙氨酸的特異性比酪氨酸提高了80,000倍。類似地,為(wei) 了能結合和轉導肽基信號(peptide-based signal)而被優(you) 化的G蛋白耦聯受體(ti) (G-protein coupled receptors),變異成了較小的遞質配體(ti) 的檢測器。

圖片來源:Pixabay
章魚的吸盤中的煙堿受體(ti) (nicotinic receptor)使研究者感到迷惑:因為(wei) 它們(men) 對乙酰膽堿不敏感。現在研究者已經認識到,這些受體(ti) 可能被多種化學感應刺激激活,而不應該局限於(yu) 它的名字的範圍內(nei) 。有關(guan) 遞質和受體(ti) 的“哪一個(ge) 先出現”的故事現在正在迅速發展,曾經困擾達爾文想象者的神經係統的神秘的起源現在變得顯而易見。
作者:John Hewitt
翻譯:蔣澤華
審校:劉宇航
引進來源:Phys.org
引進鏈接:https://phys.org/news/2021-02-brains.html
本文來自:環球科學關注【深圳科普】微信公眾號,在對話框:
回複【最新活動】,了解近期科普活動
回複【科普行】,了解最新深圳科普行活動
回複【研學營】,了解最新科普研學營
回複【科普課堂】,了解最新科普課堂
回複【科普書籍】,了解最新科普書籍
回複【團體定製】,了解最新團體定製活動
回複【科普基地】,了解深圳科普基地詳情
回複【觀鳥星空体育官网入口网站】,學習觀鳥相關科普星空体育官网入口网站
回複【博物學院】,了解更多博物學院活動詳情



 深圳市華強北街道 華新地鐵站A1出口24小時科學銀行
深圳市華強北街道 華新地鐵站A1出口24小時科學銀行










- 參加最新科普活動
- 認識科普小朋友
- 成為科學小記者




